
転写、RNAポリメラーゼ
転写
遺伝子の塩基配列をもとにして、それぞれのRNA(tRNA(トランスファーRNA)、mRNA(メッセンジャーRNA)、rRNA(リボソームRNA))を生成することを転写という。そして、この転写を行なうのがRNAポリメラーゼである。
RNAポリメラーゼ
人を含む真核生物がもつ真核細胞には、Ⅰ、Ⅱ、Ⅲの3種類のRNAポリメラーゼが存在する。ⅠはrRNAをつくり、ⅡはmRNAをつくる。ⅢはrRNAとtRNAをつくる。また、Ⅰは、細胞核内にある核小体に存在するが、ⅡとⅢは、細胞核内の液体部分である核質に存在する。
一方、細菌などの原核生物がもつ原核細胞には、1種類のRNAポリメラーゼのみ存在し、その1種類のRNAポリメラーゼで3種類のRNAをつくる。
RNAポリメラーゼは、2本鎖のDNAがもつ鎖のどちらか1本を見本として、見本となる鎖の塩基配列に結合できるRNAをつくる。
RNAの鎖にはチミン(T)がない代わりにウラシル(U)が存在する。そのため、DNA鎖のTをUに置き換える形で、見本となる鎖の塩基配列とは逆の塩基配列になる。
RNA鎖がつくられる場合も、DNA鎖をつくるときと同じように、5’末端→3’末端の方向でのみ鎖をつくることができる。
また、転写において最初につくられるRNAは、転写一次産物というRNAとしての機能をもっていないものになっている。そのため、RNAとしての機能を果たせる成熟RNAになるために、いろいろな加工(プロセシング)を受けることになる。
転写の開始、プロモーター
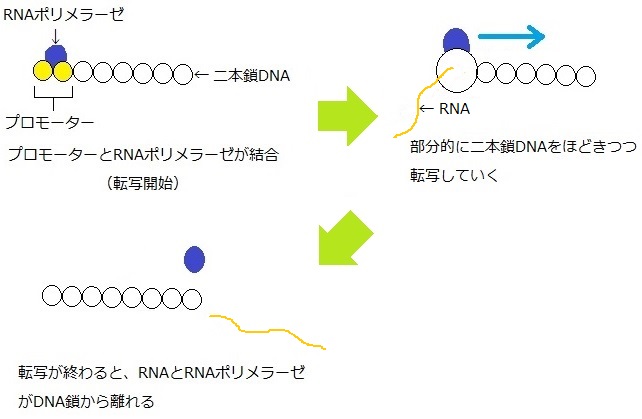
遺伝子の5’末端よりも前の位置にある、特異的な塩基配列をしたプロモーターという場所に、RNAポリメラーゼが結合するところから転写がはじまる。
RNAポリメラーゼが結合した場所にある二本鎖DNAの鎖がほどける。そして、二本鎖がほどけた部分の鎖のうち、遺伝子をもっている鎖を見本とし、その見本となる鎖が結合できる塩基配列のRNA鎖をつくっていく。二本鎖のほどけた部分は、転写が進むと同時に移動していく。
転写が終わったときは、RNAポリメラーゼとつくられたRNA鎖(転写一次産物)がDNA鎖から離れる。
転写の調節
プロモーターとなっている場所は、転写の調節にとって重要なはたらきをもつ。プロモーターを構成する基本的なものには、次のように特徴的な塩基配列の場所がある。
・TATAボックス
アデニンとチミンが多い塩基配列の場所であるTATAボックスは、5’末端側にヌクレオチド約30個分、転写開始点から離れたところにある。
・CATボックス
アデニン、チミン、シトシンが多い塩基配列の場所であるCATボックスは、5’末端側にヌクレオチド約70~80個分、転写開始点から離れたところにある。
・GCボックス
TATAボックスやCATボックスなどの塩基配列場所の代わりに、グアニンとシトシンが多い塩基配列の場所であるGCボックスが存在することもある。
・調節エレメント
プロモーターのさらに5’末端側に進んだところに、転写の調節にはたらくそれぞれの遺伝子独自の塩基配列である調節エレメントが存在する。
また、このような転写の調節にはたらく塩基配列の場所をシス・エレメントという。TATAボックスとCATボックスは、基本的シス・エレメントともいわれる。
・エンハンサー、エキソン、イントロン
遺伝子はそのほとんどが、シス・エレメントの1つであるエンハンサーによって転写を強化される。
また、遺伝子には、遺伝情報となる場所とならない場所が存在する。遺伝情報となる場所のことをエキソンといい、遺伝情報にならない場所のことをイントロンという。エキソンとイントロンは、遺伝子の中に交互に存在する。
エンハンサーは、イントロンの中に存在する場合もあり、5’末端の方向に進んだ先や、3’末端の方向に進んだ先に存在する場合もある。
転写因子(トランス因子)
それぞれシス・エレメントは、それぞれ結合したタンパク質である転写因子(トランス因子)を通してRNAポリメラーゼに影響をおよぼす。
転写因子(トランス因子)には、基本転写因子と転写調節因子がある。
基本転写因子は基本的シス・エレメント(TATAボックスやCATボックスなど)に結合する。一方、転写調節因子は、それぞれの遺伝子ごとに独自の調節エレメントと結合する。
・転写一次産物
転写で最初につくられるものを転写一次産物という。転写一次産物はRNAとして機能できないもので、RNAとして機能できる成熟RNAに変化するために、さまざまな加工(プロセシング)を受ける。
mRNAの転写一次産物の場合、5’末端でキャップ構造をつくり、3’末端でポリA鎖を付けられる。そして、イントロンを取りのぞくことであるスプライシングを受けて、成熟mRNAに変わる。
キャップ構造とポリA鎖は、どちらもmRNAを長持ちさせるために役立つものとなっている。
・非翻訳領域、翻訳領域
成熟mRNAの5’末端にあるキャップ構造の手前には、5’非翻訳領域がある。また、成熟mRNA内の3’末端のポリA鎖の手前には、3’非翻訳領域がある。非翻訳領域は、タンパク質のアミノ酸配列情報にならない場所であり、イントロンとは違う場所に存在する。
また、アミノ酸配列情報になる場所を翻訳領域という。翻訳領域は、非翻訳領域の間のエキソンが結合している部分に存在する。